Download the .pdf of the poster here. Password: OHBM2024
Introduction: Recognition of novelty constitutes a vital aspect of our ability to encode and navigate our surroundings. Though extensively studied e.g. in the context of item novelty [1], human neuroimaging studies of spatial novelty and its impact on long-term memory formation remains relatively scarce. Limited reports show that spatial novelty can boost long-term memory [e.g. 2] and is associated with distinct responses along the cortical surface and hippocampal long axis [3, 4], which requires further investigation. Here, we assessed participants during navigation of a virtual reality environment using ultra-high field 7T fMRI to chart the neural representation of spatial novelty.
Methods: A group of 56 healthy participants (20 – 37 years, M = 24 years, SD = 2.8 years, F/M ratio: 35/21) were scanned at 7T MRI (MB-BOLD, TR = 1.0 s, TE = 22.2 ms, 1.6 mm isotropic voxels) while navigating within a grassy circular environment enclosed by a wall and surrounded by landmarks (e.g. trees, buildings, mountains). Across two runs of an object location memory task, participants were asked to navigate to the locations of 6 cued objects in order to collect them and memorise their location for subsequent retrieval.
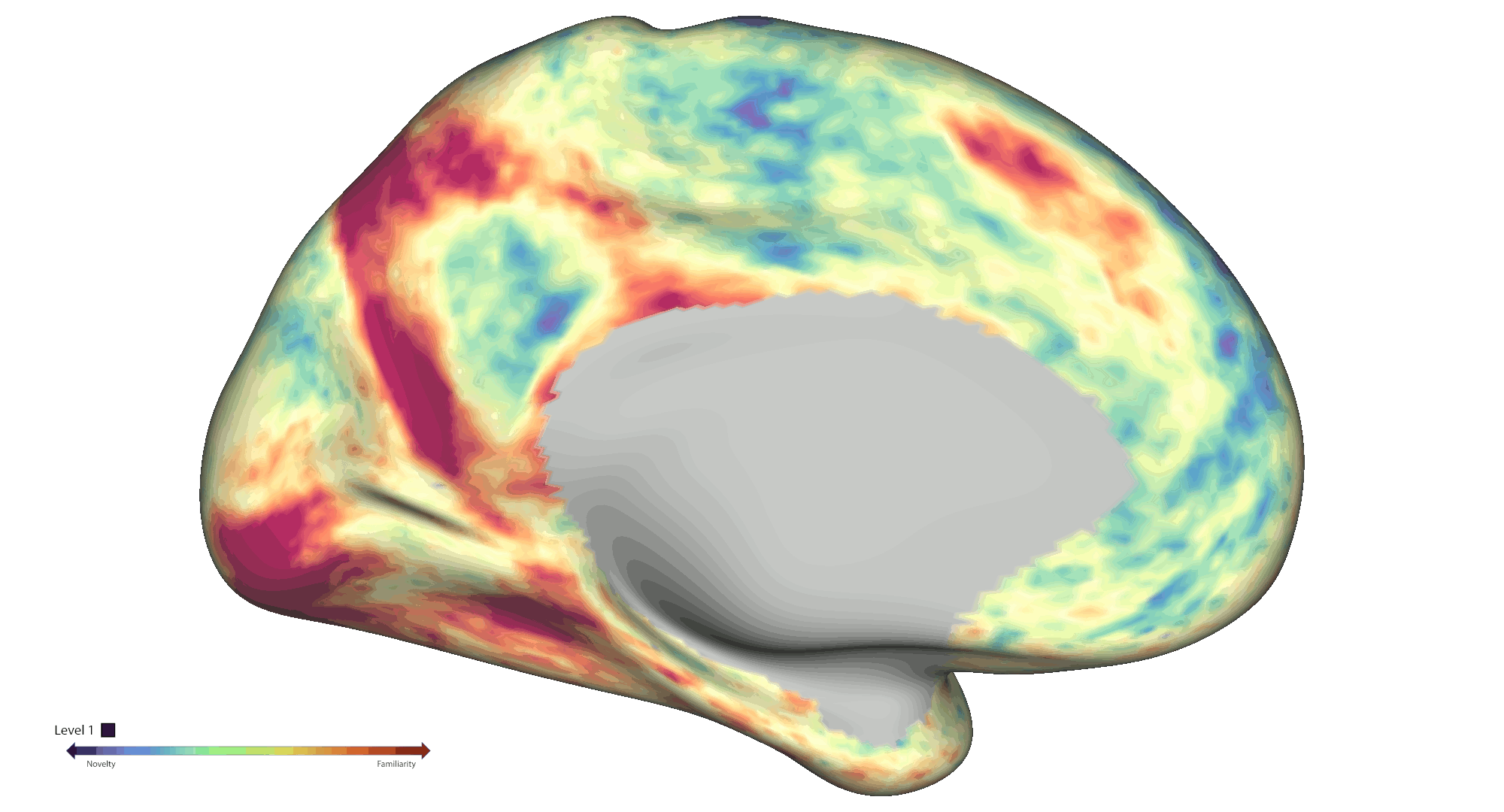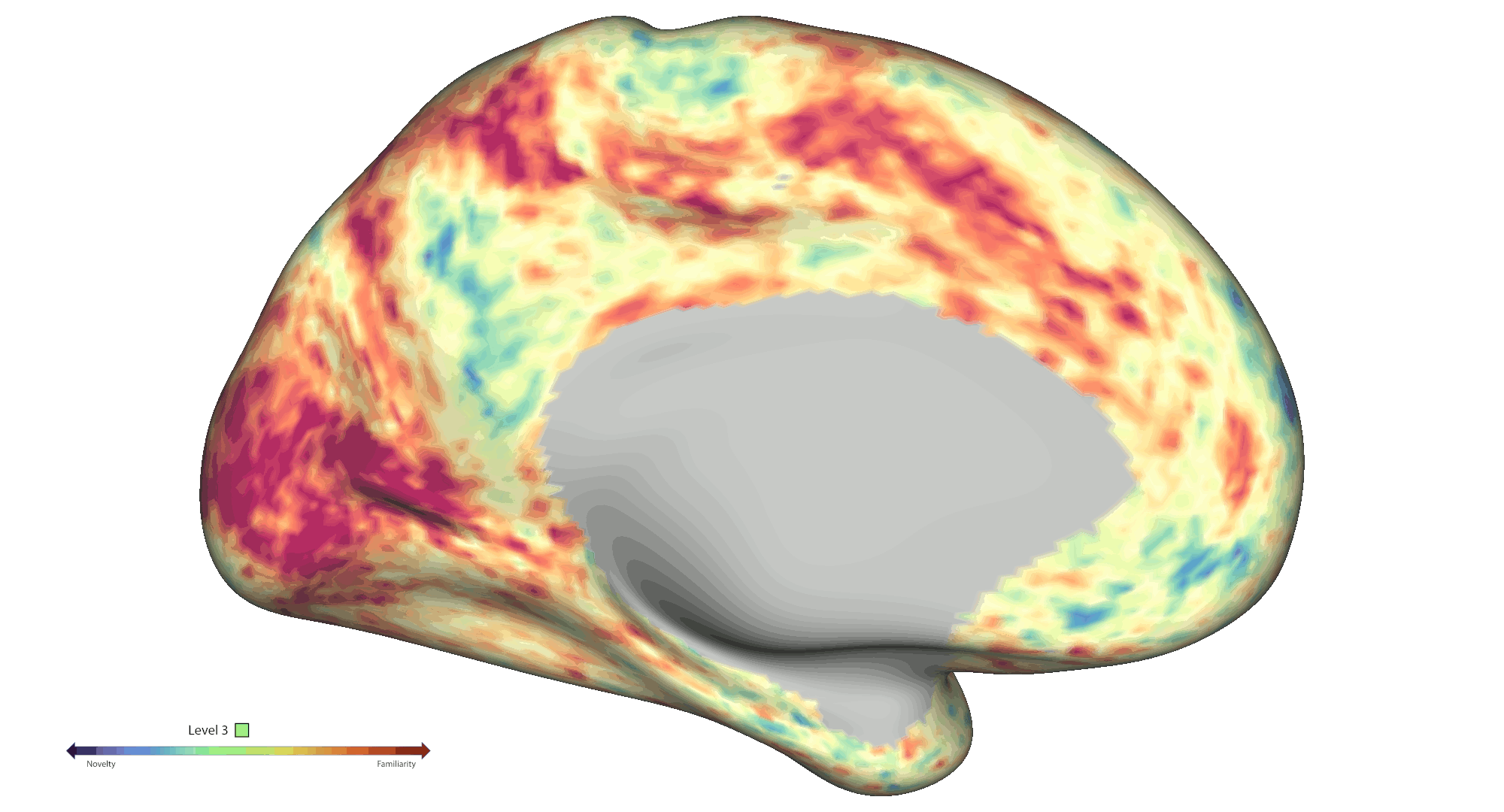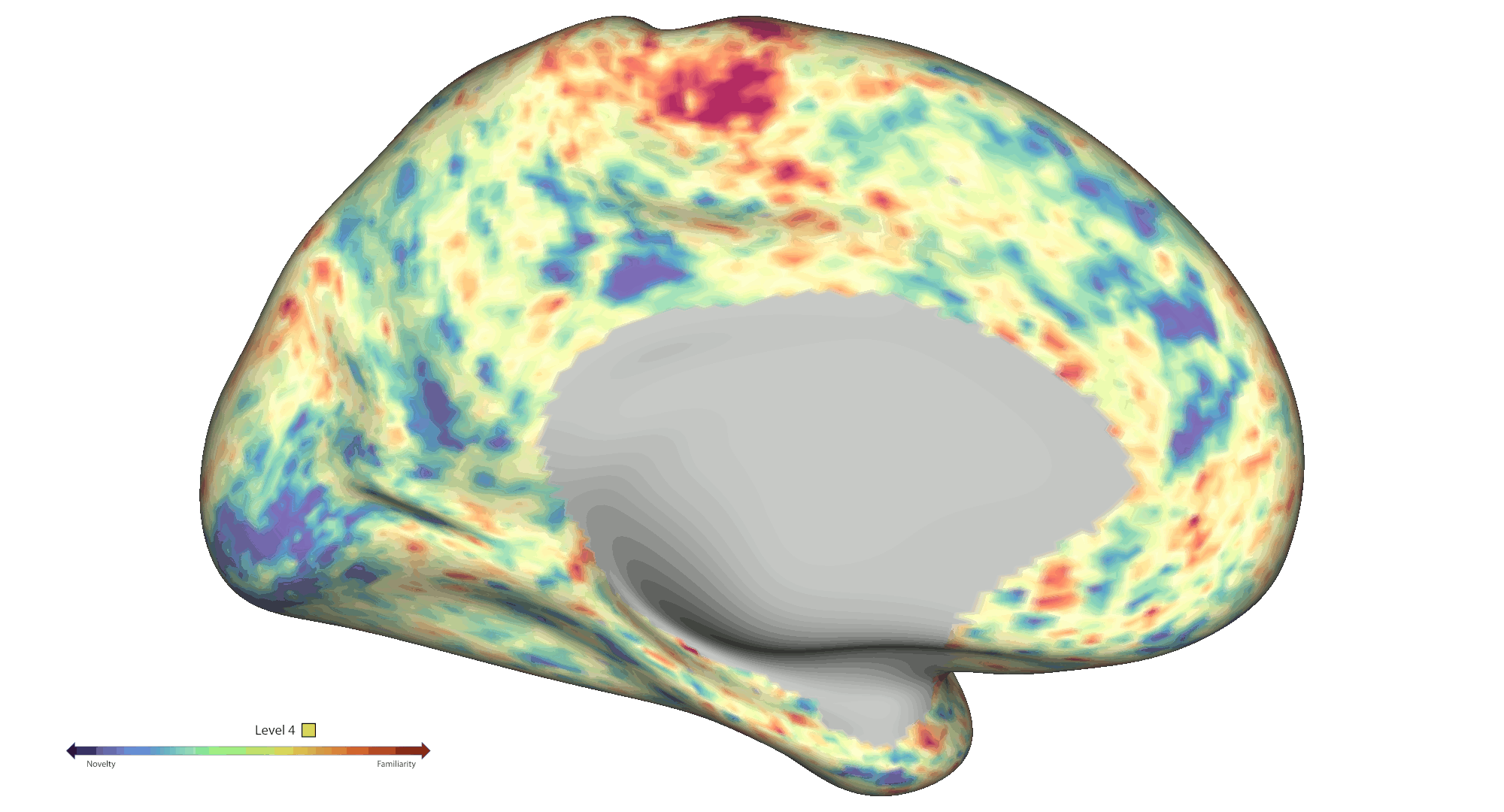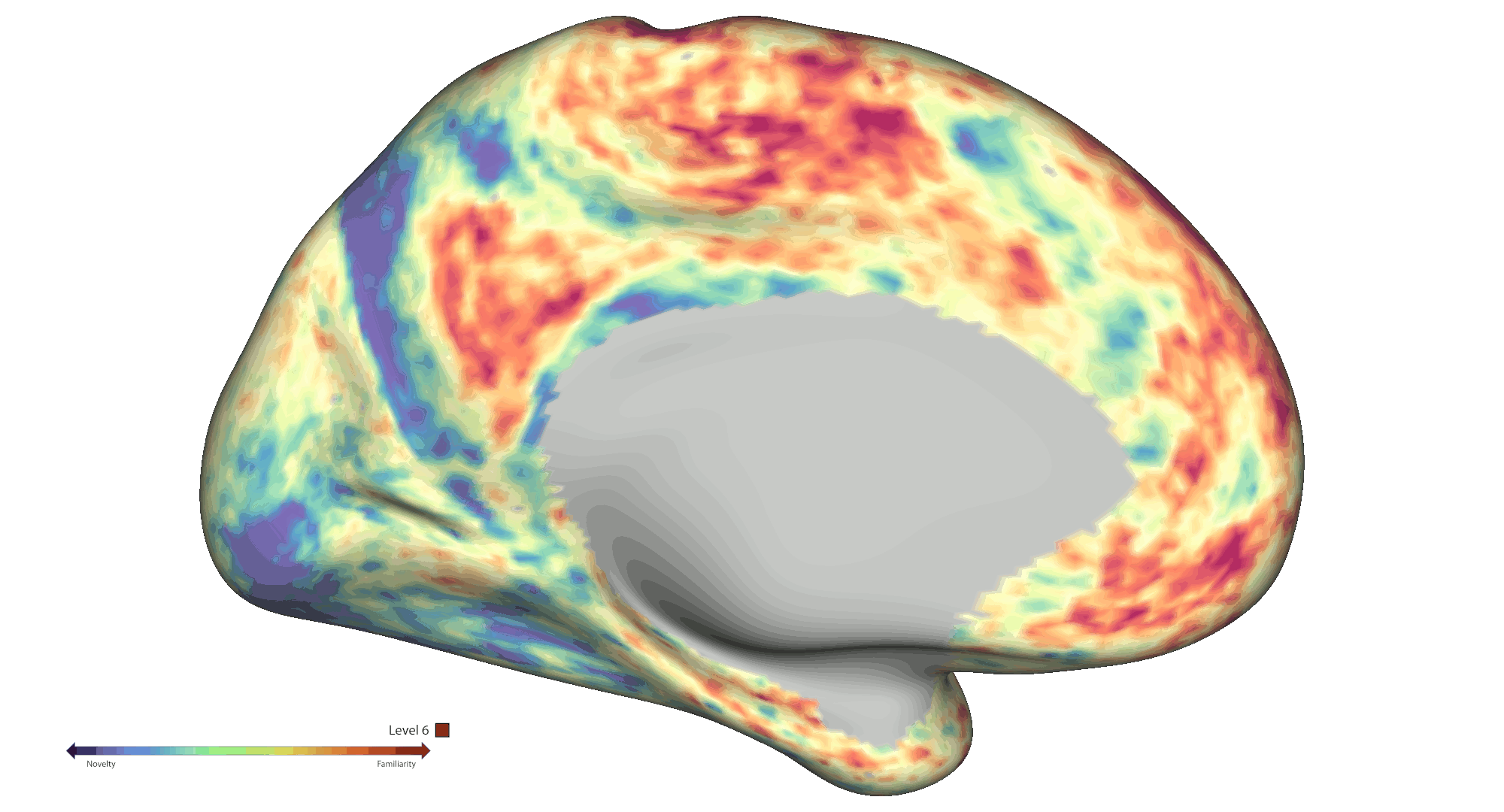
To analyse spatial novelty, the environment was divided into hexagonal sectors. Events were defined as time points that the participants spent in those sectors, which were weighted by a novelty score based on a) the number of times the sectors were visited and b) how much time passed since the last visit of the sector. The HCP-style fMRI data was pre-processed and analysed [5,6] as a GLM-based parametric modulation of events by spatial novelty using FSL FEAT routines. Statistical significance was assessed via non-parametric permutation testing based on PALM (voxel-wise cFDRp < .05). Results were projected onto cortical connectivity-based gradient space for interpretation purposes [7]. A spatial gradient analysis was also performed to further interrogate effects within the hippocampal long axis. Each voxel was assigned a value between 1 and 6 based on the novelty response [8], which were then projected on to the average shape of the hippocampi (Fig. 2a). A slope was calculated for predicting the classification based on the projection, which was finally compared to a permutation-derived null distribution.
Results: Behaviourally, novelty scores displayed an expected decrease over time due to exploration: Run 1 β = -0.48 (95 % CI [-0.56, -0.41]) and Run 2 β = -0.71 (95 % CI [-0.75, -0.66]). At the neural level, fMRI analyses across the cortical surface revealed positive modulation (greater activity for novelty) mainly centred on regions distributed across the visual and frontoparietal networks. On the other hand, negative modulation (greater activity for familiarity) was observed within somatosensory/motor and default mode areas (Fig. 1b). We also found significant negative modulation effects within the bilateral amygdala and anterior hippocampus. Notably, a significant spatial novelty-familiarity gradient was observed along the hippocampal long axis, where the posterior to anterior portions of the right hippocampus preferred to process novel to familiar sectors respectively, p < 2e-06. However, no significant spatial novelty gradient was present in the left hippocampus, p = .53 (Fig. 2a).
Conclusions: Collectively, our results highlight an extensive set of cortical and subcortical brain regions contributing to the neural representation of spatial novelty, including a novelty-familiarity gradient along the hippocampal long axis. Further work will be required to provide a more detailed mapping of the functional roles played by these regions, which will be crucial to further advance our mechanistic understanding of the novelty signature and spatial cognition in humans.
Jörn Alexander Quent1,2, Xinyu Liang1, Liangyue Song1, Yueting Su1, Kaixiang Zhuang1, Deniz Vatansever1
1Institute of Science and Technology for Brain-Inspired Intelligence, Fudan University, Shanghai, China, 200433
2MOE Frontiers Center for Brain Science, Fudan University, Shanghai, China, 200433
References:
[1] Quent, J. A., Henson, R. N. & Greve, A. A predictive account of how novelty influences declarative memory. Neurobiology of Learning and Memory 179, 107382–107382 (2021).
[2] Schomaker, J., van Bronkhorst, M. L. V. & Meeter, M. Exploring a novel environment improves motivation and promotes recall of words. Frontiers in Psychology 5, 1–6 (2014).
[3] Poppenk, J., Evensmoen, H. R., Moscovitch, M. & Nadel, L. Long-axis specialization of the human hippocampus. Trends in Cognitive Sciences 17, 230–240 (2013).
[4] Strange, B. A., Fletcher, P. C., Henson, R. N. A., Friston, K. J. & Dolan, R. J. Segregating the functions of human hippocampus. Proc. Natl. Acad. Sci. U.S.A. 96, 4034–4039 (1999).
[5] Glasser, M. F. et al. The Human Connectome Project’s neuroimaging approach. Nat Neurosci 19, 1175–1187 (2016).
[6] Ji, J. L. et al. QuNex—An integrative platform for reproducible neuroimaging analytics. Front. Neuroinform. 17, 1104508 (2023).
[7] Margulies, D. S. et al. Situating the default-mode network along a principal gradient of macroscale cortical organization. Proc. Natl. Acad. Sci. U.S.A. 113, 12574–12579 (2016).
[8] Peer, M., Ron, Y., Monsa, R. & Arzy, S. Processing of different spatial scales in the human brain. eLife 8, e47492 (2019).
Comments